Figure 7 in The Origin and Evolution of the Rattlesnake Rattle: Misdirection, Clarification, Theory, and Progress
Creators
Description
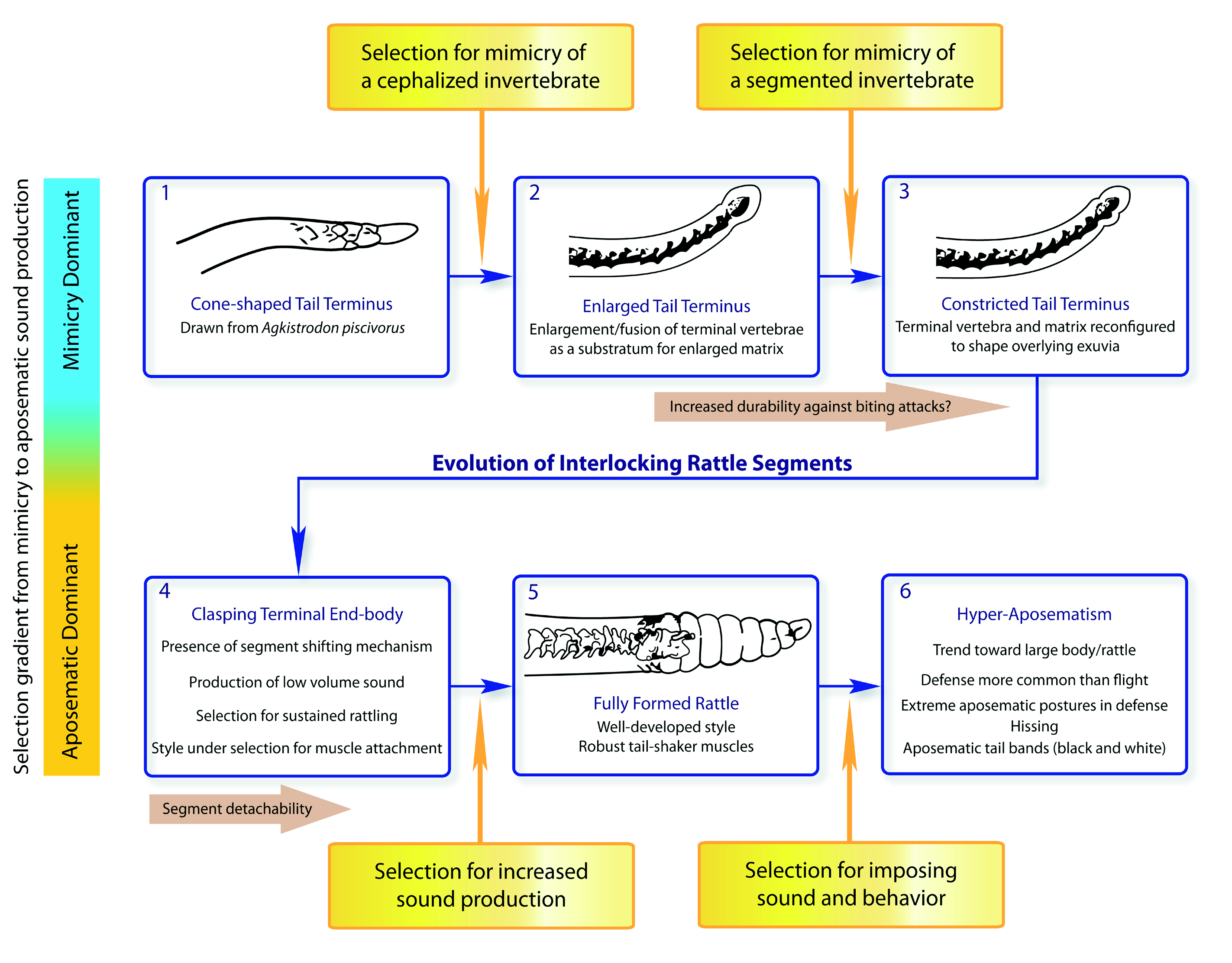
Figure 7. Unbroken sequence of evolutionary stages in rattle development illustrating the caudal luring hypothesis (CLH) of the rattle's origin. We partitioned this evolutionary sequence into six conceptual stages to emphasize possible transitions of selective influences on the incipient rattle organ system. Mimicry dominated the incipient stages (stages 1–3), which first resulted in terminal scale enlargement, followed by constriction of the end scale. Mimetic selective forces (Box 1) increased the tail's resemblance to a cephalized, segmented invertebrate, which also brought about changes in the generative tissue (matrix) and its bony substratum at the apex of the tail. In our opinion, the major evolutionary transition occurred between stages 3 and 4, when a bi- or tri-lobed end-cap, which was formerly shed during molts, gained a clasping configuration and a position shifting developmental process by which a new proximal lobe was added anterior to the old proximal lobe from the previous ecdysis (see Meik and Schuett, this volume, Rattle Evo-Devo). This innovation would have opened the door for sound production. Subsequently, with the advent of specialized rapid tail movements and changes in behavior, the aposematic role of this organ system became the dominant selective influence (stage 5). Based on multiple lines of evidence, we suggest that early rattlesnakes had small, relatively quiet rattles which could be deployed for only short durations (e.g., Sistrurus miliarius). Once aposematic sound production was achieved, certain lineages evolved hyper-aposematic, multi-modal displays (stage 6) involving increased body size (formidability), large rattles and hissing (acoustic), bold defensive postures (behavioral), starkly banded tails (black and white), larger quantities of venom per bite, and perhaps in some cases increased toxicity (Figure 8). In reference to a purely aposematic hypothesis to account for the origin of the rattle, this model fills major theoretical gaps in selective regimes present and acting on the early proto-rattle and incipient rattle. Namely, it adds stages 2 and 3, which provide an explanation for early and important morphological changes that must have required evolutionary tinkering. The transition (gradient) in selection (mimetic to aposematic) advantage appears to be complete in derived rattlesnakes (e.g., C. atrox), but is incomplete in those species that still employ caudal luring (Figures 2–4), most of which are small as adults and have limited and less dramatic aposematic repertoires (see Eberhard, 2011).
Files
figure.png
Files
(650.5 kB)
| Name | Size | Download all |
|---|---|---|
|
md5:a8fb88ee446373612e4093a4f237c78d
|
650.5 kB | Preview Download |
{kind=link}
Linked records
Additional details
Related works
- Is part of
- 10.5281/zenodo.2553648 (DOI)
- urn:lsid:plazi.org:pub:4F07FF8FFFC3271BFF5BFFF4FFA9B22E (LSID)
- https://zenodo.org/record/2553648 (URL)
- https://zenodo.org/record/2553648 (URL)